Respiration Tutorial
Click on 'Model' in the menu to the left to view this at any time. The model depicts the interrelationships between the major topics of this tutorial.
Click on "Design" in the Menu on the left.
You can select and print a copy from this page.
Design of the Model
As you read through each section below, scroll back up to the top of the page
and locate the part
of the model being described.
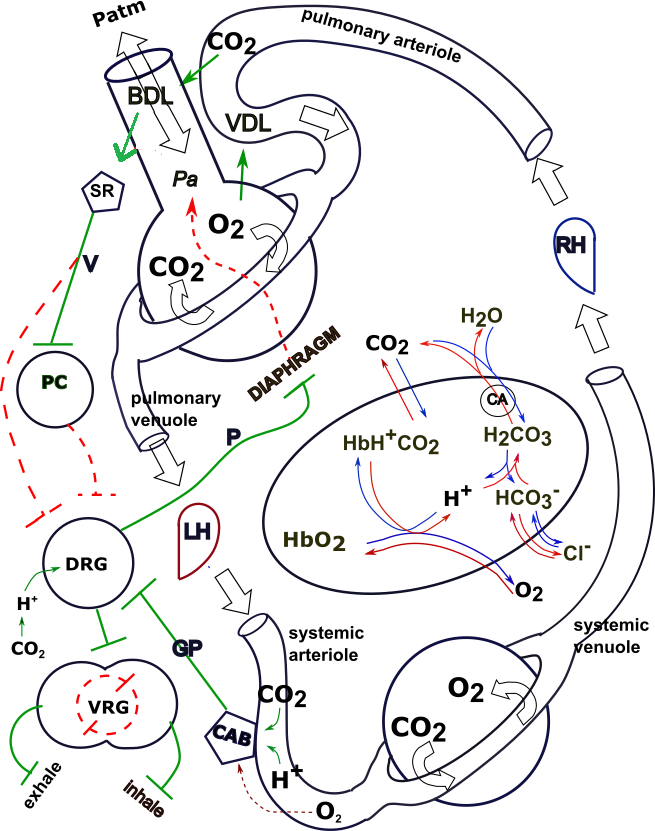
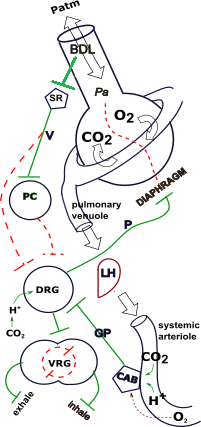
The flask at the upper left represents an alveolus and a respiratory passageway. The donut encircling it represents pulmonary capillaries. A pulmonary arteriole is at the top of the model carrying blood to the alveolus and air passage; a pulmonary venuole below the alveolus carries blood to the left heart (LH).
The circle at the bottom of the model represents a body cell; the encircling donut represents systemic capillaries. Blood is brought to these capillaries via systemic arterioles and blood is carried away by systemic venuoles that eventually lead to the right heart (RH) .
The oval in the center represents a red blood cell. Various chemical reactions are shown to occur inside the cell. The circles and small pentagons seen along the left side of the model represent various neural pathways that control breathing.
Symbolism
Throughout the model nerves are represented as both solid and dashed lines with small perpendicular lines at their ends. Solid green nerves indicate that these will cause a stimulatory response of their target; dashed red nerves indicate inhibitory responses.
Solid and dashed red and green arrows indicate non-neural relationships; solid green arrows represent direct relationships while dashed red arrows indicate inverse relationships.
The thin red and blue arrows within and around the oval RBC indicate chemical reactions.
Respiration
Respiration is the exchange of oxygen and carbon dioxide across capillary walls throughout the body. Exchange of these gases with the outside environment is external respiration and takes place across the walls of pulmonary capillaries. Exchange of these gasses between the blood and cells throughout the body is internal respiration and takes place across the walls of systemic capillaries.

Systemic Gas Exchange / Internal Respiration
The insert shows a body cells encircled by systemic capillaries. Blood enters via systemic arterioles (left) and leaves through systemic venuoles (right). The insert shows carbon dioxide entering the blood and oxygen leaving the blood.
Carbon Dioxide
Carbon dioxide is a waste product of metabolism and is constantly produced in cells throughout the body. This maintains a concentration gradient guaranteeing that carbon dioxide will diffuse from systemic cells into the capillaries as indicated by the arrow. As a result the concentration of carbon dioxide in the blood increases from 40 mm Hg to 45 mm Hg.
Oxygen
Having passed through the lungs, blood entering systemic capillaries is rich in oxygen (104 mm Hg). As a result, oxygen diffuses from the blood into the cells as shown by the arrow. When blood leaves the body cells, via the systemic venuoles, its concentration has dropped to 40 mm Hg.

Pulmonary Gas Exchange / External Respiration
The inset shows an alveolus and air passage of the lungs. Blood enters the pulmonary capillaries (donut) via pulmonary arterioles (right) and leaves via pulmonary venuoles (left). Atmospheric oxygen within the alveolus diffuses into the blood as shown by the arrow; carbon dioxide leaves the blood (arrow) and enters the alveolus. The double-headed arrow represents the inward and outward flow of air (ventilation).
Carbon Dioxide
Carbon dioxide is at extremely low levels in the atmosphere (0.3 mm Hg). Blood entering the pulmonary capillaries has picked up carbon dioxide from body cells and has a partial pressure of 45 mm Hg. Therefore, this gas diffuses out of the blood (arrow) and into the alveoli. When the blood leaves the pulmonary capillaries its concentration has decreased to 40 mm Hg. This decrease seems small but it is only measuring the carbon dioxide dissolved in the plasma; much more carbon dioxide is in the form of bicarbonate ions and bound within red blood cells as described in the next section.
Oxygen
Blood entering the pulmonary capillaries has an oxygen concentration of 40 mm Hg. The concentration of atmospheric oxygen is 160 mm Hg so oxygen diffuses from the alveoli into the blood. Oxygen in the blood leaving the lungs has increased to 104 mm Hg.
Transport of Respiratory Gases
Cell membranes are completely permeable to the passage of carbon dioxide and oxygen. On diffusing into the red blood cell these gases interact with other compounds. Once beyond the capillaries no further gain or loss of these gases can occur and the compounds involved reach an equilibrium.
While in the systemic capillaries, the RBCs are subjected to increasing levels of carbon dioxide and decreasing levels of oxygen. The opposite is the case in the pulmonary capillaries. The reactions are the same but in reverse directions in these two locations. In the model the blue arrows represent the direction of the reactions occurring in the systemic capillaries; the red arrows represent the direction in the pulmonary capillaries.
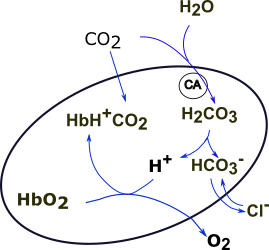
Chemical Reactions in Systemic Capillaries--Blue Arrows
The enzyme carbonic anhydrase (CA in circle) is found inside red blood cells. This enzyme catalyzes the combination of carbon dioxide (CO2) and water (H2O) into carbonic acid (H2CO3). Once formed within the RBC a small proportion of carbonic acid molecules ionize to produce hydrogen ions (H+) and bicarbonate ions (HCO3-).
The RBC membrane is also freely permeable to bicarbonate ions (HCO3-). These ions diffuse into the plasma along their concentration gradient in exchange for chloride ions (Cl-). This is called the 'chloride shift.'
The hydrogen ions (H+) in the RBC are buffered, that is, they are not allowed to diffuse into the plasma. Instead, they bind to hemoglobin (Hb) but only after they displace oxygen (O2) from that compound. Hemoglobin, when it binds to hydrogen ions, is said to be 'reduced.' It is in this form that hemoglobin also binds with carbon dioxide to form carbaminohemoglobin (HbH+CO2).
Carbon dioxide is transported in three forms. The smallest amount is in the form of dissolved carbon dioxide located in the plasma. There is somewhat more carbon dioxide in the form of carbaminohemoglobin confined to the RBC. The majority is in the form of bicarbonate ions that equilibrate across the RBC membrane.
Chemical Reactions in Pulmonary Capillaries--Red Arrows
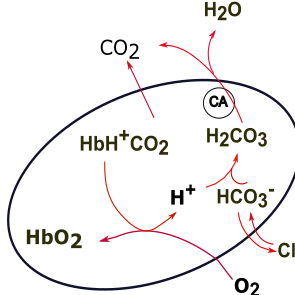
As oxygen (O2) enters the RBC it interacts with carbaminohemoglobin after displacing hydrogen ions (H+) from it. This also causes any bound carbon dioxide to dislodge. In normal situations hemoglobin becomes almost 100% saturated with oxygen. In this form it is called oxyhemoglobin (HbO2).
The 'freed' hydrogen ions (H+) combine with bicarbonate ions (HCO3-) to form carbonic acid (H2CO3). This reduces the bicarbonate ions within the RBC causing more to diffuse in from the plasma. This, in turn, causes a chloride shift from the RBC into the plasma.
The additional carbonic acid (H2CO3) within the RBC is converted to carbonic acid and water by carbonic anhydrase (CA in circle). The increased carbon dioxide in the plasma (45 mm Hg) diffuses into the alveoli when blood traverses pulmonary capillaries.
Ventilation & Perfusion
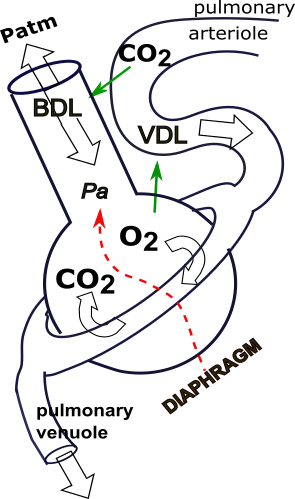
Ventilation
Ventilation (breathing) moves air into and out of the lungs. As the diaphragm contracts it enlarges the thoracic cavity which, in turn, drops the pressure in this space. This causes the pliable lungs to expand into this space which, in turn, decreases the pressure within the lungs. There now exists a pressure gradient between the atmospheric air (Patm) and alveolar air (Pa) causing inward air flow (inhalation).
Alveolar pressure (Pa) is controlled by the activity of the diaphragm. When the diaphragm contracts, causing the thoracic cavity to expand, the alveolar pressure is reduced--an inverse relationship represented by the dashed red arrow. This establishes a pressure gradient where alveolar pressure is lower than atmospheric pressure (Patm) and inhalation occurs. When the diaphragm relaxes, allowing the thoracic cavity to recoil to a smaller volume, the alveolar pressure increases reversing the pressure gradient ... air flows from the alveoli to the atmosphere (exhalation) along this new pressure gradient.
Ventilation-Perfusion Coupling
Perfusion is blood flow within an organ and a unique anatomical design exists between pulmonary arterioles and bronchioles leading to alveoli. As shown in the insert, pulmonary arterioles travel parallel and adjacent to these bronchioles. This close physical contact allows compounds in the blood to affect bronchioles as well as allowing compounds in the bronchiolar air to affect the arterioles. This is ventilation-perfusion coupling.
The respiratory bronchioles (neck of the flask) contain smooth muscle tissue that controls their diameter. If these muscles relax, bronchodilation (BDL) occurs promoting air flow. Bronchoconstriction is the opposite; narrower airways present more resistance to air flow through them. The close contact between these bronchioles and arterioles permits the diffusion of gases between blood and air.
When carbon dioxide concentration in the blood increases the adjacent bronchiolar smooth muscles relax and the bronchioles dilate (BDL). This direct relationship ... increased blood carbon dioxide and increased bronchodilation ... is represented by the (solid green arrow).
This coupling also exists in the reverse direction. Oxygen in alveolar air affects a smooth muscle in the arterioles causing vasodilation (VDL). The solid green arrow indicates this direct relationship between alveolar oxygen concentration and blood flow through these arterioles. Increased alveolar oxygen concentration increases arteriolar vasodilation to insure good blood flow (perfusion) surrounding alveoli with good oxygen levels.
Respiratory Center
The job of the respiratory system is to maintain systemic arterial oxygen levels at 104 mm Hg and carbon dioxide at 40 mm Hg. However, during exertion, systemic venous blood levels of carbon dioxide increase while oxygen levels decrease rapidly; the rate and depth of breathing must increase to re-establish normal values in a timely manner.
The brainstem contains three paired groupings of neurons that control breathing by regulating the rate and depth of ventilation. The dorsal respiratory group sets an intrinsic basic rhythm of breathing. The pneumotaxic center can change the rate of this basic rhythm. The ventral respiratory group comes into play when there is need for increased depth of each breath such as during exercise.
Some some texts refer to an apneustic center though its physical existence has not been proven. It is characterized by regular deep inspirations followed by a slight pause before an inadequate expiration; usually occurs due to brain injury. It is not shown in this model.
Quiet Breathing (eupnea)
The dorsal respiratory group (DG), often referred to as the inspiratory center, sets the basic rhythm of ventilation. During quiet breathing this consists of inhalation for 2 seconds followed by passive expiration for 3 seconds (12 breaths per minute); this rate is referred to as eupnea.
Inspiratory neurons travel in the phrenic nerve (P) from the DRG to target the diaphragm and external intercostal muscles that contract to expand the thoracic cavity. Action potentials transmitted along this nerve are weak to begin with but gradually become stronger so that each inhalation is gradual and not a gasp; this gradually-increasing pattern is referred to as the inspiratory ramp. The action potentials stop abruptly after 2 seconds allowing passive relaxation of the thoracic cavity. The reason for this inherent oscillation is complex and not fully understood.
Active Breathing (hyperventilation)
Many factors play a role in modifying the basic rhythm set by the DRG. Here we will consider changes in lung volume and changes in respiratory gases.
Lung Volume
There are stretch receptors (SR) embedded in the smooth-muscle walls of bronchiolar smooth muscle that become increasingly activated during bronchodilation (BDL). Their neurons (dashed red line) target the DRG to stop inhalation thus preventing over-inflation beyond ~800 ml. This phenomenon is called the Hering-Breuer reflex.
There are other types of 'stretch receptors' located mainly in the upper respiratory passages. These respond to a variety of stimuli and are more appropriately called 'irritant receptors'. Neurons leave these receptors via the vagus nerve (V) (solid green line) to target the normally passive pneumotaxic center (PC). This center responds by sending impulses (dashed red line) to the DRG decreasing the inhalation component of the basic rhythm. Since inhalation is shortened there are more breaths per minute.
Respiratory Gases
The model illustrates the ventral respiratory group (VRG) as a fused double circle. Though it contains neurons that target muscles involved with exhalation (exhale) and inhalation (inhale) though its primary outflow is to expiratory muscles. These are the internal intercostals and the abdominal muscles. The VRG is not active during normal quiet breathing. The dashed red circle within the VRG represents a neural mechanism that insures inhalation and exhalation do not occur simultaneously. The DRG receives central and peripheral input that causes it to communicate with the VRG (solid green line).
Central Chemoreceptors
Central receptors (not shown) on the surface of the medulla respond to changes in carbon dioxide levels. This is represented at the left of the DRG. There is no central response to oxygen levels in the blood.
Capillaries in the brain are not permeable to hydrogen ions but readily let carbon dioxide penetrate the blood-brain barrier. Carbonic anhydrase in cerebrospinal fluid converts carbon dioxide to carbonic acid that releases hydrogen ions. The solid green arrows indicate the DRG responds to increases in carbon dioxide via the accompanying increase in hydrogen ions. This response causes stimulation of the neurons leading to the VRG to include forced expiration in the normally passive breathing pattern.
Peripheral Chemoreceptors
The peripheral chemoreceptors are paired neuron groups located bilaterally in the carotid and aortic bodies (CAB). They respond strongly to decreased (dashed red arrow) blood oxygen levels and show a weaker response to increased (solid green arrows) carbon dioxide and hydrogen ion levels. They are connected to the dorsal respiratory group (DRG) by the glossopharyngeal (GP) nerve (solid green line). This mechanism is the major response to blood oxygen levels while the central receptors supply the main response to blood carbon dioxide levels.