Excretion Tutorial
Click on 'Model' in the menu to the left to select and print a copy of the model .
Click on "Design" to begin the tutorial.
Overview
Plasma containing both wastes and nutrients passes into the nephrons of the kidneys where the nutrients are returned to the blood and the wastes left behind in a fluid called 'filtrate.' This filtrate is refined on its way to becoming 'urine.' In addition to removing waste from blood plasma the kidneys regulate the degree of hydration of the body.
Scroll down for a description of the model's design.
As you read through each section below, scroll back up to the top of the page
and locate the part
of the model being described.
Design of the Model
Nephron
This structure is outlined in black and begins near the upper left corner of the model. The cup-shaped Bowman's capsule is the first portion followed by the proximal convoluted tubule shown as a horizontal scalloped region.
Decending down the right side of the model is a straight region representing the decending limb of Henle. At the lower right corner of the map this bends back on itself to become the ascending limb of Henle. The walls of the ascending limb are thick to represent its impermeability to water.
The following region--also impermeable to water--bends to the left and is scalloped and horizontal; this is the early distal convoluted tubule. It is shown folding back on itself with its walls drawn as a thick but broken line that symbolizes its permeability to water is variable. This region is the late distal convoluted tubule.
The final region of the nephron is the collecting duct shown as a vertical extension of the late distal convoluted tubule. It's permeability to water is also variable.
Blood Supply
The structure outlined in red represents the blood supply. Pressing into Bowman's capsule is the glomerulus. The region arching up and over Bowman's capsule is the efferent arteriole.
The next region that runs horizontally beneath the proximal convoluted tubule represents the peritubular capillaries. At the right the vessel is drawn straight and vertical to represent the descending limb of the vasa recta. At the bottom of the model it folds back upon itself and becomes the ascending limb of the vasa recta.
The next region that turns to the left to become horizontal represents, once more, the peritubular capillaries. Next there is a gap in this vascular system indicating we are leaving the kidneys. The separate U-shaped region represents the extrarenal vascular system
The new start of the vascular system represents a return to the kidneys as the afferent arteriole. As it passes upward it contacts the distal convoluted tubule forming the juxtaglomerular apparatus. This structure consists of two parts. On the vascular side is a region called the juxtaglomerular cells, JGC and on the nephron side is a region called the macula densa, MD.
Central Nervous System
At the center of the left margin of the model are two circles that represent the vasoconstrictor center, VC and the vasodilator center, VD.
Glomerular Filtration
The glomerulus is a capillary bed enclosed by Bowman's capsule. The filtration membrane' separating the blood plasma and the filtrate in the capsule is impermeable to large plasma proteins but permits the passage of water, electrolytes and small molecules. The filtrate formed passes into the proximal convoluted tubule (PCT).

Vascular Component
The afferent arteriole supplies the glomerulus with blood. Blood leaves the glomerulus via the efferent arteriole that quickly gives rise to the peritubular capillaries.
Filtrate Formation
There are three factors that control the volume of filtrate (F) produced from blood plasma.
Blood Hydrostatic Pressure
The pressure against the inside of the capillary wall is called blood hydrostatic pressure (HPb). An increase in blood pressure (BP) will increase the hydrostatic pressure as indicated by the solid arrow.
The block arrow labeled F represents the filtrate entering the capsule. An increase in blood hydrostatic pressure (HPb) will increase the volume of filtrate (F) formed as indicated by the solid arrow between these two symbols.
Blood Osmotic Pressure
Because the 'filtration membrane' is impermeable to large plasma proteins they accumulate in the remaining plasma. This increases the blood osmotic pressure (OP) causing water to return to the plasma from the filtrate. Thus, as osmotic pressure increases the net volume of filtrate decreases as indicated by the dashed arrow.
Capsular Hydrostatic Pressure
Filtrate accumulation inside Bowman's capsule establishes a capsular hydrostatic pressure (HPc) that opposes the formation of additional filtrate. This is represented by the dashed arrow pointing back to the block filtration arrow.
Proximal Convoluted Tubule
This is the first region of the nephron the filtrate enters after its formation in the glomerulus. It is surrounded by peritubular capillaries. The primary activities that occur here are 'reabsorption' of specific components and production of the hormone erythropoietin. This hormone stimulates bone marrow to produce red blood cells. Several specific components are retrieved from the filtrate and water follows the osmotic gradient this creates.
Erythropoietin
Erythropoietin production is represented by the pink block arrow. The capillary endothelial cells in this region are sensitive to the oxygen concentration in the blood and release this hormone into that blood when oxygen levels are low. When this hormone circulates through the bone marrow is stimulates the production of new red blood cells thus increasing the oxygen-carrying capacity of the blood.
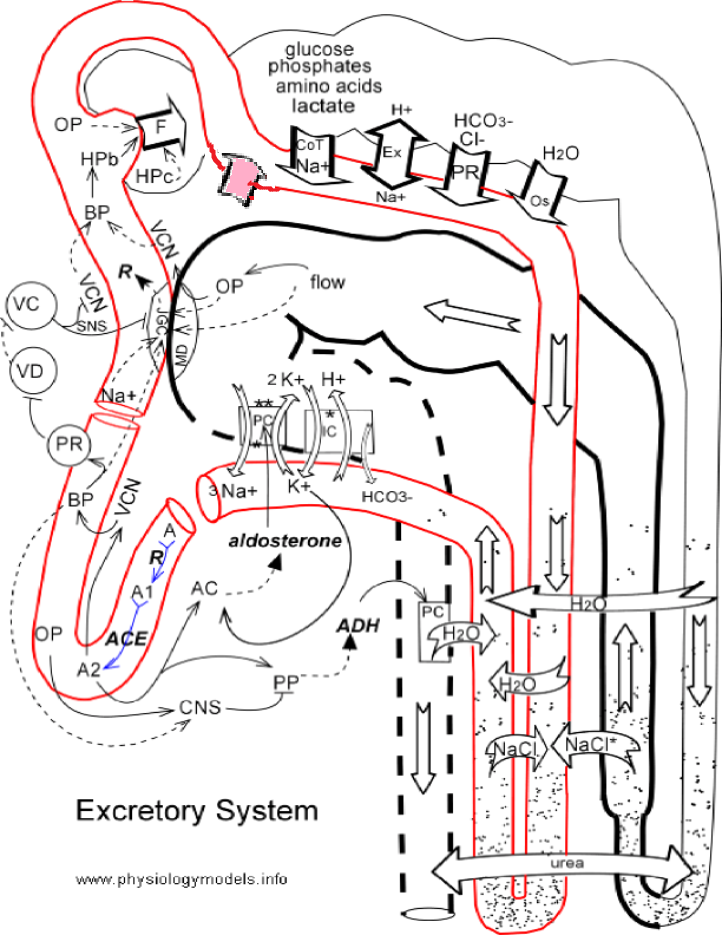
Reabsorption
Cotransport
There are specific active transport pumps in the proximal tubule cells that simultaneously move (i.e., cotransport) sodium (Na+) with glucose, phosphates, amino acids, or lactate from the filtrate into the peritubular capillaries. This is represented by the block arrow labeled CoT Na+.
Exchange
These cells also move hydrogen ions (H+) from the blood to the filtrate in exchange for sodium ions (Na+). This is represented by the double-headed block arrow.
Passive Reabsorption
Negative ions such as chloride (Cl-) and bicarbonate (HCO3-) diffuse from the filtrate into the blood along the electrical gradient established by the active movement of the positive ions mentioned earlier. This is represented by the block arrow labeled PR.
Osmosis
The movement of the above mentioned solutes establishes an osmotic gradient that causes water (H2O) to diffuse from the filtrate and into the blood. This movement of water is referred to as 'obligatory' because it must follow the osmotic gradient that is established in this location. This is represented by the blocked arrow labeled Os.
Counter-Current Mechanism
The mechanisms shown here are traditionally called the 'counter-current multiplier' and the 'counter-current exchanger'. The first takes place in the region of the nephron called Henle's loop; the second occurs in a region of the peritubular capillary bed called the 'vasa recta'. Both are involved in establishing an osmotic gradient throughout this region.

Henle's Loop
The portion of the nephron called 'Henle's loop' is shown in the right half of the insert. It consists of a descending limb that has a down arrow in it and an ascending limb with an up arrow in it. The ascending limb has a thick and a thin segment. The dots represent various solutes that become more concentrated in both limbs toward the bottom of the loop.
The thick walls of the ascending limb indicates that this region is impermeable to water.
Vasa Recta
This capillary bed is shown in red at the left of the insert. It also consists of a descending limb and an ascending limb identified by the arrows in each. Again, the dots represent various solutes that are more concentrated at the bottom of the loop.
Counter-Currents
Counter-currents exist when fluids flow in opposite directions in parallel and adjacent tubes. The two limbs of Henle's loop are a counter-current and so are two limbs of the vasa recta. The same is true of the ascending limb of Henle and the descending vasa recta. Though not drawn adjacent in the insert, the descending limb of Henle is also counter-current with the ascending limb of the vasa recta;
Counter-Current Exchanger
Examination of both limbs of the vasa recta shows the concentration of solutes (number of dots) is the same at any horizontal level. However, imagine the fluid flowing through the vasa recta for a short distance then stopping. Now compare the imaginary concentration of solutes at any level and they will not be the same. At any level the solute concentration in the descending limb will be less than in the ascending limb! But, because both limbs are freely permeable, sodium chloride will diffuse from the ascending into the descending while water will diffuse from the descending to the ascending...see block arrows. When equilibrium is reached both limbs will, once again, have the same concentration of water and solutes. Water is exchanged for sodium chloride...the counter-current exchange mechanism.
Counter-Current Multiplier
The above described counter-current exchanger would not exist if there were not some mechanism to initially make the vasa recta more concentrated at the bottom of the loop. This is accomplished by the loop of Henle.
The ascending limb of Henle and the early distal tubule are impermeable to water as indicated by their thick wall. These regions actively transports sodium chloride (NaCl) out of the filtrate and into the surroundings. (The asterisk (*) in the block arrow indicates that a more complex mechanism is involved but the net effect is that only NaCl is moved out of the filtrate.)
The NaCl diffuses into the descending limb of the vasa recta...block arrow. Any that might diffuse into the descending limb of Henle will only get pumped back out when it enters the ascending limb so this is not shown in the model. It will not diffuse into the ascending vasa recta because that fluid already has a high concentration of NaCl. This is the mechanism that 'multiplies' the concentration of NaCl in the descending vasa recta making the counter-current exchanger possible!
Osmosis
Both blood and filtrate descending into their respective loops have low solute concentrations. Both flow beside upcoming columns having higher solute concentrations. As they move past one another, water from the down-flowing fluid columns will diffuse into the more concentrated up-flowing columns. However, only the up-flowing vasa recta is permeable to water meaning all is returned to the blood and none to the filtrate...see both block arrows. This also insures that a high solute concentration will be maintained at the bottom of both loops.
Urea
Urea passively diffuses among and between the lower portions of both the blood and the filtrate loops as indicated by the large block arrow spanning this region. The result is that half the urea is excreted and half kept in the body.
Distal Tubule & Collecting Duct
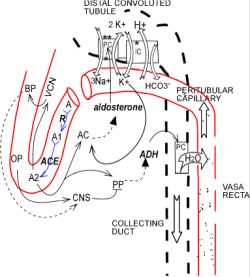
This region of the nephron is under the control of two hormones: aldosterone and antidiuretic hormone (ADH). Potassium, acid-base, and water balance are controlled here.
Intercalated Cells
This type cell is found in the late distal tubule and is represented by the IC box. Its main function is acid-base balance.
The basolateral border of this cell is freely permeable to hydrogen and potassium. As blood becomes more acidic, hydrogen ions (H+) equilibrate with this cell and electrostatically send potassium ions (K+) from it into the blood as indicated by the block arrows. The hydrogen ions are secreted into the filtrate by hydrogen/potassium exchange pumps in its luminal border(*).
'New' bicarbonate ions (HCO3-) are formed within this cell by the action of carbonic anhydrase. Only the basolateral border is permeable to these ions and they diffuse into the blood as indicated by the block arrow.
Principal Cells
This common type of cell is represented by the two PC boxes. It is the major type of cell in the late distal tubule and the collecting duct. Its main function is to regulate potassium and water balance.
The hormone aldosterone affects this cell by stimulating (solid arrow) it to produce additional sodium/potassium pumps in its basolateral border. This pump moves 3 sodium ions (Na+) from the filtrate into the peritubular capillaries in exchange for 2 potassium ions (K+) as indicated by the two curved block arrows. The hormone also increases the number of potassium channels in the luminal border (**) to facilitate the diffusion of this ion from the cell and into the filtrate.
The hormone ADH also affects this cell by stimulating it to produce water channels in its otherwise impermeable luminal border; this is indicated by the thick broken wall in this region of the nephron. In the late distal tubule the simultaneous presence of both hormones allows water to 'follow salt' into the peritubular capillaries (not indicated in the model). In the collecting duct ADH allows water to diffuse (block arrow) from the filtrate into the hypertonic blood in the ascending vasa recta.
Aldosterone
Two conditions stimulate the adrenal cortex (AC) to secrete (solid-head dashed arrow) this hormone.
Potassium Level
An increase in blood potassium (K+) stimulates (solid arrow) the adrenal cortex (AC) to secrete (solid-headed dashed arrow) aldosterone. The result will be to lower blood potassium due to increased activity of the principal cells.
Angiotensin II
This hormone is formed when the enzyme renin (R) is present in the blood. Renin converts the inactive compound angiotensinogen (A) into angiotensin I (A1) as indicated by the reaction arrow. The enzyme angiotensin converting enzyme (ACE), bound to the walls of pulmonary capillaries, catalyzes the conversion of A1 into the more potent compound angiotensin II (A2) as indicated by the second reaction arrow. This hormone stimulates the adrenal cortex (AC) to secrete (solid-headed dashed arrow) aldosterone. It also causes (solid arrow) generalized vasoconstriction (VCN) which increases (solid arrow) blood pressure (BP).
Antidiuretic Hormone
This hormone is secreted by the posterior pituitary (PP) as indicated by the solid-headed dashed arrow. ADH affects the principal cells by increasing their permeability to water as indicated by the thick broken walls of the late distal convoluted tubule and the collecting duct. This permits the reabsorption of water (block arrow) into the vasa recta. The secretion of this hormone is controlled by two mechanisms.
Angiotensin II
Renin (R) is released when blood pressure is low. As a result angiotensin II (A2) is formed as discussed above. This hormone stimulates the release of ADH as indicated by the solid-headed dashed arrow.
Central Nervous System
When the osmotic pressure (OP) of the blood is high the central nervous system will stimulate (solid arrow) the posterior pituitary (PP) to release ADH. The blunt headed line represents this neural connection. The effect will be to increase water reabsorption thus reducing blood osmotic pressure.
High blood pressure (BP) will inhibit (dashed arrow) the central nervous system from signaling the posterior pituitary to release ADH. The effect will be to reduce water reabsorption, reduce blood volume and reduce blood pressure.
Justaglomerular Apparatus
This is a region where, in each nephron, the distal convoluted tubule touches the afferent arteriole. The juxtaglomerular cells are part of the afferent arteriole and the macula densa is part of the distal tubule. Renin is secreted from the JG cells and is important in regulating systemic blood pressure. This region also regulates local blood pressure to control the rate of filtrate formation in its glomerulus-- a mechanism called 'tubuloglomerular feedback'.

Systemic Blood Pressure
Renin (R) is secreted (solid-headed dashed arrow) into the blood by the juxtaglomerular cells (JGC). The end effect is to lower blood pressure. This enzyme catalyzes the first step in the formation of angiotensin II as described in the Angiotensin II section of the previous mini-tutorial. There are several mechanisms that control renin secretion.
Sodium Concentration
There is an inverse relationship (dashed arrow) between sodium (Na+) concentration and the secretion (solid head on arrow) of renin. An increase in sodium causes a decrease in renin. One result of decreased renin is decreased sodium reabsorption due to decreased aldosterone levels.
Blood Pressure
The relationship with renal blood pressure (BP) is also inverse (dashed arrow). An increase in renal blood pressure caused a decrease in renin.
Neural Controls
An increase in systemic blood pressure (BP) activates (solid arrow) pressure receptors (PR) in the large arteries near the heart. These send stimulatory signals (blunt-headed solid line) to the vasodilator center (VD) (VD circle). The VD center sends inhibitory signals (blunt-headed dashed line) to the vasoconstrictor center (VC) (VC circle).
Without input from the VD center the fibers from the VC center send a constant stream of stimulatory signals (solid arrow) to the juxtaglomerular cells (JGC) and the vascular smooth muscle (VCN) of the afferent arteriole. This causes a base line secretion (solid-headed dashed arrow) of renin (R) and a basal state of constriction in these these arterioles. With input from the VD center the activity of the VC center decreases (blunt-headed dashed line) to reduce renin secretion and vasoconstriction in the afferent arteriole.
Tubuloglomerular Feedback
High glomerular blood pressure cause a high rate of filtration (F). The greater the volume of filtrate formed the more rapidly it passes through the nephron. This rapid flow allows some solutes to pass through the proximal tubule and Henle's loop without being reabsorbed. The result is increased rate of flow and increased osmotic pressure (OP) in the distal convoluted tubule of this nephron.
The macula densa (MD) on the tubule side of the juxtaglomerular apparatus is sensitive to the osmotic pressure (OP) and/or the rate of flow of the filtrate. These cells seem to produce a yet unknown compound that inhibits dashed arrows the JGC from producing renin in this nephron.
It is also speculated that increased osmotic pressure (OP) stimulates the production of some compound that directly affects (solid arrow) the vascular smooth muscle (VCN) of the adjacent afferent arteriole. The result is decreased (dashed arrow) glomerular blood pressure leading to a decreased rate of filtrate (F) production in this nephron.